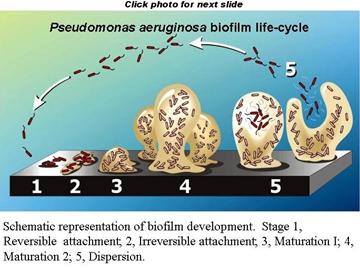
Biofilm formation is
a developmental process in which bacteria undergo a regulated lifestyle switch
from a nomadic unicellular state to a sedentary multicellular state where
subsequent growth results in structured communities and cellular
differentiation. Results of prior work by many groups allow the construction of
a hypothetical developmental model for biofilm formation that can be
generalized for many different bacterial species. This model can be adjusted to
fit either of two general modes of unicellular lifestyle: nonmotile and motile.
In the case of
non-motile species, when conditions are propitious for biofilm
formation, individual bacteria appear to increase the expression of adhesins on
their outer surface, i.e., they increase their “stickiness”. This increased
stickiness promotes both cell-cell adherence and cell-surface adherence when
these bacteria encounter a surface. For example, in the case of
some strains of staphylococcal species, surface-expressed proteins, including Bap,
promote cellcell interaction and contribute to the extracellular matrix. Many other species,
both nonmotile and motile, harbor homologs of Bap. A unifying feature of these
large extracellular proteins is the presence of repeated domains.
At the level of the bap gene, these repeats
have been shown to be recombinogenic, resulting in the production of proteins
of variable length within a biofilm population. Yet, the
significance of this variability in the size of Bap proteins within a biofilm
remains unknown. Nonmotile species also produce exopolysaccharides (EPS) that
form an integral part of the extracellular matrix. One example of this is the
PIA or PNAG EPS, produced by the gene products of the ica operon of
staphylococcal species. Thus, in nonmotile bacteria, changes in cell surface
proteins, along with the production of EPS, play a critical role in the initiation
of biofilm formation.
In the case of motile
species, when conditions favor biofilm formation, individual bacteria
localize to a surface and initiate a dramatic lifestyle switch. Motility is
lost and bacteria begin to produce an extracellular matrix that holds the cells
together. For a number of motile organisms, the dominant role for flagella in initiation
of biofilm formation is to provide motility as flagella-minus and paralyzed flagella
mutants are comparably defective in biofilm formation. In fact, in Listeria
monocytogenes supplying exogenous cell movement directed toward the surface via
centrifugation restores wild type levels of initial surface adhesion to
nonmotile mutants. In these cases, it appears that motility
is the driving force that overcomes repulsive forces between the bacteria and
the surface. Initial encounters with a surface usually lead to transient
adherence. This transient adherence can result in either a stable surface
association, and a subsequent switch to biofilm development, or in a return to
planktonic existence.
No comments:
Post a Comment